|
|||
|
|||
Νευρωνική απώλεια των ιππόκαμπων στην κροταφική επιληψία
ΖΑΦΕΙΡΟΠΟΥΛΟΣ Α., ΞΑΦΕΝΙΑΣ Δ.
Α' Νευρολογική Κλινική Α.Π.Θ. Νοσοκομείο ΑΧΕΠΑ,
Εργαστήριο Νευροπαθολογίας
Περίληψη
Ποικίλες παθολογοανατομικές μελέτες ιππόκαμπου που προέρχονταν από ασθενείς οι οποίοι έπασχαν από κροταφική επιληψία, ενισχύουν την άποψη ότι η νευρωνική απώλεια του ιππόκαμπου συνδέεται άμεσα με τη σύνθετη εστιακή επιληψία, διαπιστώνοντας ότι στο 90% των ασθενών με εστιακές κρίσεις (αύρες) που προηγούνταν των επιληπτικών κρίσεων του κροταφικού λοβού εμφάνιζαν άλλοτε άλλου βαθμού απώλεια των νευρώνων του ιππόκαμπου. Περιοχές με κύρια επικράτηση νευρωνικής απώλειας αναφέρονται στην οδοντωτή ταινία, στο Αμμώνειο κέρας, στο προϋπόθεμα και στο υπόθεμα ενώ το μεταϋπόθεμα και η ιπποκάμπεια έλικα δεν παρουσίαζαν παραδόξως καμιά κυτταρική μεταβολή.
Η ερμηνεία αυτής της σημαντικής νευρωνικής απώλειας στην T.L.E. είναι ασαφής. Ίσως οι επιληπτικές κρίσεις να προκαλούν την ιπποκάμπεια σκλήρυνση αλλά ίσως να συμβαίνει και το αντίθετο, η ιπποκάμπεια σκλήρυνση να προκαλεί τις επιληπτικές κρίσεις. Συνήθως στην έναρξη της της ιπποκάμπειας σκλήρυνσης -όρος που υποδηλώνει τη νευρωνική απώλεια στην TLE- ο ένας ιππόκαμπος προσβάλλεται περισσότερο από τον άλλο. Αυτή η παρατήρηση της ετερόπλευρης ευπάθειας των ιππόκαμπων αποκλείει την άποψη μιας σφαιρικής τοξικής ή μεταβολικής αιτιοπαθογένειας. Αν η νευρωνική απώλεια οδηγεί σε επιληπτικές κρίσεις, θα πρέπει ένας κριτικής σημασίας υποπληθυσμός νευρώνων να υφίσταται νέκρωση στην TLE, καθώς γενικευμένη διάσπαρτη νευρωνική απώλεια του ιππόκαμπου, που παρατηρείται στην ισχαιμία, την υποξεία, τη νόσο Alzheimer, τις εγκεφαλίτιδες ή τις δηλητηριάσεις από CO δεν επαρκεί για την πρόκληση κρίσεων. Έτσι, φαίνεται πλέον πιθανό το γεγονός ότι οι κρίσεις προκαλούν την παραπάνω νευρωνική απώλεια του ιππόκαμπου και με τη σειρά της αυτή η νευρωνική απώλεια ίσως να οδηγεί στις υποτροπιάζουσες κρίσεις.
Πρόσφατες μελέτες υποστηρίζουν την άποψη ότι η νευρωνική απώλεια προηγείται της ανάπτυξης της επιληψίας και είναι η κύρια αιτία της έναρξης των παθολογικών διεργασιών από τους νευρώνες για την απελευθέρωση αυτής. Στην ιπποκάμπεια σκλήρυνση, η διαπίστωση της αδυναμίας των βρυωδών κυττάρων να φτάσουν μέχρι τα καλαθιοφόρα κύτταρα περιορίζει την παλίνδρομη καταστολή που απελευθερώνουν αυτά πάνω στα κοκκιώδη ή πυραμοειδή κύτταρα, θέτοντας τα θεμέλια για τη δημιουργία της υπόθεσης των λανθανόντων καλαθιοφόρων κυττάρων, όπου οι ενδιάμεσοι κατασταλτικοί GABAεργικοί νευρώνες του επιληπτικού ιππόκαμπου, λόγω μερικής αποσύνδεσής τους από τις διεγερτικές προσυναπτικές ίνες βρίσκονται σε "λανθάνουσα", δηλαδή ανενεργό κατάσταση. Η διαπίστωση της θεωρίας των λανθανόντων καλαθιοφόρων κυττάρων έχει συντελέσει στην ανεύρεση και εφαρμογή σημαντικών θεραπευτικών παρεμβάσεων με παράγοντες που ενισχύουν τη GABAεργική καταστολή.
Συνοψίζοντας το σύνολο των παραπάνω αναφορών, τόσο σε ανθρώπινο όσο και σε πειραματικό υλικό ζώων, συμπεραίνεται, ότι η επιληπτογένεση είναι το αποτέλεσμα της εκλεκτικής απώλειας ορισμένων νευρώνων, οι οποίοι φαίνεται να διαδραματίζουν ένα σημαντικό ρόλο στη μείωση του ουδού για απελευθέρωση της επιληπτικής δραστηριότητας σε έναν κατά τα άλλα φυσιολογικό εγκέφαλο ή σε εγκέφαλο που εμφανίζει μια γενετική προδιάθεση για κρίσεις. Η αιτία για την έναρξη της νευρωνικής απώλειας είναι άγνωστη, αλλά είναι σχεδόν βέβαιο, ότι στα πρώϊμα στάδια της επιληπτογόνου διεργασίας, ορισμένοι νευρώνες κλειδιά, εμφανίζουν δυσλειτουργία, η οποία οδηγεί σε επιληπτικές κρίσεις και ιπποκάμπεια σκλήρυνση. Οι επαναλαμβανόμενες κρίσεις με τη σειρά τους, ίσως να επιβαρύνουν τη διεργασία της σκλήρυνσης στον ήδη επιληπτικό εγκέφαλο. Έτσι, μολονότι η μη ειδική κυτταρική απώλεια των νευρώνων του ιππόκαμπου που μπορεί να παρατηρηθεί σε ποικίλες κλινικές οντότητες, δεν σχετίζεται με επιληψία, η ειδική κυτταρική απώλεια ενός νευρωνικού πληθυσμού του ιππόκαμπου, η οποία συνδέεται άμεσα με επιληψία, μπορεί να προκαλεί παθολογική ευερεθιστότητα των διεγερτικών κυκλωμάτων, που οδηγούν σε επαναλαμβανόμενες επιληπτικές κρίσεις.
Λέξεις κλειδιά: Κροταφική επιληψία, νευρωνική απώλεια, ιπποκάμπεια σκλήρυνση, καλαθιοφόρα κύτταρα, βρυώδη κύτταρα.
Παθολογοανατομία της κροταφικής επιληψίας
Πρόσφατες μελέτες ιππόκαμπων, που προέρχονταν από ασθενείς, οι οποίοι έπασχαν από κροταφική επιληψία (TLE), αναφέρονται στην περιγραφή των παθολογοανατομικών αλλοιώσεων οι οποίες παρατηρούνται σε αυτοψικό υλικό. Η πιο παλαιά αναφορά, η οποία περιγράφει τις παθολογοανατομικές αλλοιώσεις του ιππόκαμπου, προέρχονταν από ασθενείς, οι οποίοι έπασχαν από TLE, ήταν μια μελέτη αδρών ιστοτεμαχίων αυτοψικού υλικού, όπου παρατηρήθηκαν για πρώτη φορά "ορατές" (ψηλαφητές) αλλοιώσεις στους 8 ιππόκαμπους από τους 14 μελετηθέντες εγκεφάλους ασθενών που έπασχαν από εστιακή επιληψία1. Σε μια μεταγενέστερη, το 1880, μελέτη εγκεφάλων, οι οποίοι προέρχονταν από νεκροτομικό υλικό επιληπτικών ασθενών, διαπιστώθηκε ότι η ασύμμετρη ατροφία των ιππόκαμπων σχετίζονταν άμεσα με την επιληψία2. Μεταγενέστερες ιστολογικές μελέτες, ενισχύουν την άποψη ότι η νευρωνική απώλεια των ιππόκαμπων συνδέεται άμεσα με τη σύνθετη εστιακή επιληψία (Complex Partial Epilepsy-CPE)3-5. Μία συσχέτιση μεταξύ της CPE και της νευρωνικής απώλειας του ιππόκαμπου για πρώτη φορά υποστηρίχτηκε από τον Stauder το 1936 και στα 1959 αποκαλύφθηκε ότι το 90% των ασθενών με εστιακές κρίσεις (αύρες), που προηγούνταν των επιληπτικών κρίσεων του κροταφικού λοβού εμφάνιζαν άλλοτε άλλου βαθμού απώλεια των νευρώνων του ιππόκαμπου4.Τα προηγούμενα ευρήματα επαναβεβαιώθηκαν από τους Margerison and Corsellis το 19665-8.
Ποικίλοι όροι έχουν χρησιμοποιηθεί για να περιγράψουν τη νευρωνική απώλεια του ιππόκαμπου που συχνά παρατηρείται στην TLE, όπως ιπποκάμπειος σκλήρυνση (hippocampal sclerosis), ή εσωτερική κροταφική σκλήρυνση (mesial temporal sclerosis), καθώς και σκλήρυνση του Αμμωνείου κέρατος (Ammon's horn sclerosis). Οι όροι αυτοί υποδηλώνουν τη νευρωνική απώλεια και την αντικατάσταση αυτής από κύτταρα της νευρογλοίας στον επιληπτικό εγκέφαλο. Ο όρος εσωτερική κροταφική σκλήρυνση χρησιμοποιήθηκε κυρίως για να περιγράψει μια νευρωνική απώλεια των εξωιπποκαμπικών δομών, όπως του αμυγδαλοειδή πυρήνα και της παραϊπποκάμπειας έλικας. Ο όρος σκλήρυνση του Αμμωνείου κέρατος φαίνεται να είναι ατυχής, καθώς εκτός απ' αυτό προσβάλλεται και η οδοντωτή έλικα. Από τη στιγμή που τα ειδικά παθολογοανατομικά ευρήματα της TLE έχουν διασαφηνιστεί, οι όροι ιπποκάμπειος και εσωτερική κροταφική σκλήρυνση, αδιαφιλονίκητα μπορούν να χρησιμοποιηθούν, καθώς καλύπτουν πλήρως τα πιο πάνω ευρήματα.
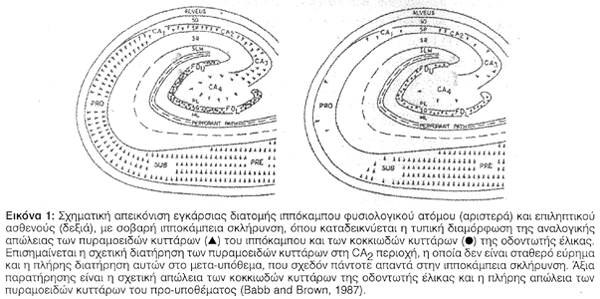
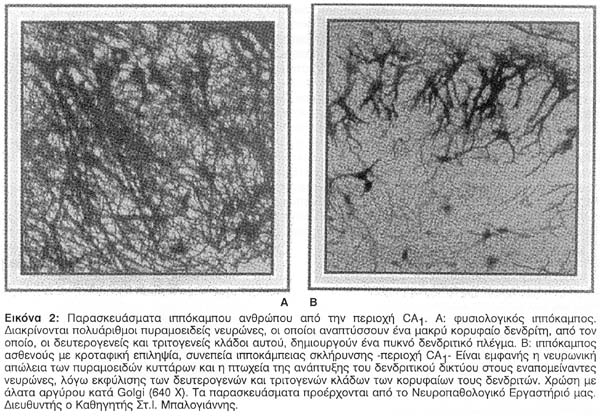
Λεπτομερείς μελέτες των παθολογοανατομικών ευρημάτων του ιππόκαμπου αποκαλύπτουν μία ευρείας έκτασης ανομοιογένεια των περιοχών που εμφανίζουν νευρωνική απώλεια στην TLE. Περιοχές με κύρια επικράτηση νευρωνικής απώλειας αναφέρονται στην οδοντωτή ταινία (fascial dentata), στο Αμμώνειο κέρας, στο προϋπόθεμα (prosubiculum) και στο υπόθεμα (subiculum) αλλά όχι στο μεταϋπόθεμα (presubiculum) ή την ιπποκάμπεια έλικα (hippocampal gyrum)9,10. H Εικόνα 1 περιγράφει μiα σχηματική αναπαράσταση της σχετικής έκτασης της κυτταρικής απώλειας στην ιπποκάμπεια σκλήρυνση, σύμφωνα με μία μελέτη των Babb και Brown το 19874. Από τις περιοχές, που υπέστησαν νευρωνική καταστροφή φαίνεται ότι η CA1 είναι η πλέον έκδηλα επηρεασμένη (Εικόνα 2), και ακολουθείται από τη CA3 και τη CA2, ενώ η CA4και η οδοντωτή έλικα φαίνονται ελάχιστα επηρεασμένες9. Ορισμένοι ερευνητές έχουν περιγράψει μια σχετική κυτταρική πτωχεία στη CA2 περιοχή8. Στο 60-65% των ασθενών με TLE, η ιπποκάμπειος σκλήρυνση εμφανίζεται ως το μοναδικό παθολογοανατομικό εύρημα. Το υπόλοιπο 35-40% των ασθενών αποκαλύπτει άλλου τύπου παθολογοανατομικές αλλοιώσεις, όπως μεταστατικούς όγκους και άλλες εξεργασίες, για παράδειγμα αμαρτώματα, που συνυπάρχουν με ιπποκάμπεια νευρωνική απώλεια4. Σ' αυτές τις περιπτώσεις η κυτταρική απώλεια του ιππόκαμπου δεν είναι τόσο σοβαρή, όσο στην αμιγή ιπποκάμπειο σκλήρυνση, η οποία σχετίζεται με την κρυπτογενή (ιδιοπαθή) TLE11.Το πως και το εάν οι πιο πάνω εξεργασίες και οι διαμαρτίες περί τη διάπλαση προκαλούν ιπποκάμπεια διαταραχή παραμένει αδιευκρίνιστο και ελάχιστα μελετημένο.
Η αιτιολόγηση αυτής της σημαντικής νευρωνικής απώλειας στην TLE είναι ασαφής. ΄Ισως επιληπτικές κρίσεις να προκαλούν την ιπποκάμπειο σκλήρυνση αλλά ίσως να συμβαίνει και το αντίθετο δηλαδή η ιπποκάμπειος σκλήρυνση να προκαλεί τις επιληπτικές κρίσεις. Όταν η ιπποκάμπειος σκλήρυνση εκδηλωθεί, συνήθως ο ένας ιππόκαμπος προσβάλλεται περισσότερο από τον άλλο. Αυτή η ετερόπλευρη ευπάθεια των ιππόκαμπων υποδηλώνει ότι, όταν ξεκινάει η διεργασία της επιληπτογένεσης αυτή επηρεάζει πολύ περισσότερο τον έναν ιππόκαμπο, απ' ότι τον άλλο, διαφωνώντας με τις απόψεις μιας σφαιρικής τοξικής ή μεταβολικής αιτιοπαθογένειας, η οποία, όπως είναι γνωστό υπεισέρχεται σε μια γενικευμένη κρίση, που εκδηλώνεται από λήψη φαρμάκων, κατάσταση υποξείας ή από τοξίνες του περιβάλλοντος. Από την άλλη πλευρά μία εστιακή κρίση, η οποία ξεκινάει μέσα από τον ιππόκαμπο ή από γειτονικές περιοχές αυτού μπορεί να προκαλεί αυτή την ασύμμετρη ιπποκάμπειο σκλήρυνση. Εάν αυτή η νευρωνική απώλεια οδηγεί σε επιληπτικές κρίσεις θα πρέπει ένας κριτικής σημασίας υποπληθυσμός νευρώνων να υφίσταται νέκρωση στην TLE, καθώς γενικευμένη διάσπαρτη νευρωνική απώλεια του ιππόκαμπου, όπως αυτή που παρατηρείται στην ισχαιμία, στην υποξεία, στη νόσο του Alzheimer, στις εγκεφαλίτιδες ή σε δηλητηρίαση από CO, δεν επαρκεί για την πρόκληση κρίσεων4. Πιο πιθανό λοιπόν φαίνεται ότι οι κρίσεις προκαλούν την παραπάνω νευρωνική απώλεια του ιππόκαμπου, και με τη σειρά της αυτή η νευρωνική απώλεια ίσως να οδηγεί σε υποτροπιάζουσες κρίσεις (επιληψία).
Κλινικές μελέτες καταλήγουν στο συμπέρασμα ότι η νευρωνική απώλεια, ιδιαίτερα όταν διαπιστώνεται σε μικρή ηλικία, μπορεί να δίνει το έναυσμα για την έναρξη της ιπποκάμπειας σκλήρυνσης. Μία μελέτη υποστηρίζει στενή σχέση μεταξύ μιας σοβαρής παιδικής νόσου και ιπποκάμπειας σκλήρυνσης σε ενήλικες ασθενείς με TLE9. Μία άλλη μελέτη προβαίνει στην υπόθεση ότι η ιπποκάμπεια σκλήρυνση είναι πλέον συχνή στους ασθενείς με έναρξη των κρίσεων πριν από τον τέταρτο χρόνο ζωής τους12. Σύνθετες μυοκλονικές κρίσεις της βρεφικής ηλικίας έχουν ενοχοποιηθεί ως μία αιτία ιπποκάμπειας σκλήρυνσης13,14, καθώς επίσης και οι πρώιμοι παιδικοί σπασμοί15, μολονότι σημαντική συσχέτιση μεταξύ των σύνθετων μυοκλονικών κρίσεων και της TLE δεν έχει αναφερθεί σε άλλη μελέτη16.Μία πρόσφατη μελέτη σε ιππόκαμπους, που προέρχονταν από 26 ασθενείς με TLE αποκάλυψε ότι το 50% των νευρώνων με παράπλευρα αξονικά κλωνία, που εμφανίζουν εκτροπή ανήκουν σε ασθενείς με πρώιμη εμφάνιση κρίσεων17.Μολονότι φαίνεται ότι σε ορισμένες κακώσεις πρώϊμης ηλικίας μπορεί να συντελούν την έναρξη των διεργασιών της ιπποκάμπειας σκλήρυνσης, άλλοι αιτιογενετικοί παράγοντες, που εμπλέκονται στους αρχικούς παθογενετικούς μηχανισμούς του κυτταρικού θανάτου, όπως η ενδογενής κωδικοποίηση του κυτταρικού θανάτου, καθώς και ο ρόλος των αναπτυξιακών επιδράσεων στην ιπποκάμπεια σκλήρυνση παραμένει ασαφής.
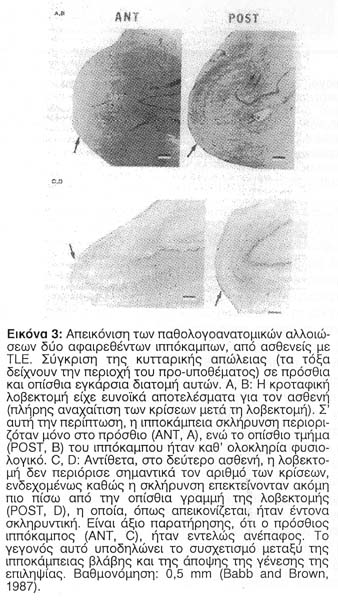
Πρόσφατες μελέτες σε εγκεφαλικό ιστό ανθρώπου υποστηρίζουν την άποψη, ότι η νευρωνική απώλεια προηγείται της ανάπτυξης της επιληψίας. Αυτοί οι ερευνητές εξέτασαν την κυτταρική πυκνότητα διαφόρων περιοχών σε 45 χειρουργικά αφαιρεθέντες επιληπτικούς κροταφικούς λοβούς και τους σύγκριναν με 4 κροταφικούς λοβούς από νεκροψικό υλικό, ως ομάδα ελέγχου. Διαπίστωσαν τότε μια κλιμάκωση της κυτταρικής απώλειας με επιδείνωση αυτής της κλιμάκωσης από τις πρόσθιες προς τις οπίσθιες περιοχές του ιππόκαμπου. Ιδιαίτερα σ' εκείνους τους ασθενείς που παρατηρήθηκε μεγαλύτερη κυτταρική απώλεια στις πρόσθιες ιπποκάμπειες περιοχές η μετεγχειρητική ανταπόκριση ήταν σαφώς καλύτερη από ό,τι σε εκείνους, που εμφάνιζαν οποιαδήποτε άλλου χαρακτήρα κατανομή της κυτταρικής απώλειας. Επίσης απέδειξαν ότι η κυτταρική απώλεια, η οποία εντοπιζόταν στις πρόσθιες ιπποκάμπειες περιοχές συνδεόταν σταθερά στους ασθενείς με ΗΕΓ ευρήματα έναρξης κρίσεων από τις πρόσθιες περιοχές του ιππόκαμπου, σε αντίθεση με εκείνους τους ασθενείς, που παρουσίαζαν ΗΕΓ ευρήματα με έναρξη των κρίσεων οι οποίες προέρχονταν από ευρύτερες περιοχές του ιππόκαμπου και υποδήλωναν μία περισσότερο ευρείας έκτασης νευρωνική απώλεια. Ο Babb και οι συνεργάτες του το 1990 υποθέτουν ότι η σκλήρυνση μπορεί μερικώς να σχετίζεται με την αιτία της ιπποκάμπειας επιληψίας ή να προκαλεί το έναυσμα έναρξης παθολογικών διεργασιών από νευρώνες για την απελευθέρωση αυτής. Η Εικόνα 3 περιγράφει την τοπογραφική σχέση της κυτταρικής απώλειας στον ιππόκαμπο του ανθρώπου9.
Μελέτες σε ζώα υποστηρίζουν ότι η κυτταρική απώλεια είναι η αιτία έναρξης της ανάπτυξης της ιπποκάμπειας σκλήρυνσης. Σε επίμυες, επαναλαμβανόμενα ηλεκτρικά ερεθίσματα στις ίνες της διάτρητης οδού για διάρκεια 24 ωρών18, προκαλούν υπερευερεθιστότητα και μείωση τόσο της ορθόδρομης, όσο και της παλίνδρομης καταστολής στα κοκκιώδη κύτταρα της CA1 περιοχής της οδοντωτής έλικας, η οποία εμμένει για χρονικό διάστημα πλέον των 24 ωρών. Αυτή η απόσυρση της καταστολής παρατηρήθηκε μόνο σ' εκείνους τους ιππόκαμπους, που παρουσίαζαν απώλεια των βρυωδών κυττάρων της πύλης, καθώς και των κυττάρων τα οποία περιέχουν τη σωματοστατίνη και το νευροπτιτίδιο Υ, αλλαγές που έχουν περιγραφεί και σε καταστάσεις TLE στον άνθρωπο19,20. Ανεπηρέαστη έκφραση της κατασταλτικής δραστηριότητα της CA1 περιοχής του ιππόκαμπου διαπιστώθηκε όταν χορηγήθηκε ερέθισμα στην ετερόπλευρη περιοχή των ινών της διάτρητης οδού, γεγονός που οδηγεί στην υπόθεση ότι ο μηχανισμός της ιπποκάμπειας καταστολής ήταν παρόν μεν αλλά ανενεργός. Η αδυναμία των βρυωδών κυττάρων να φτάσουν μέχρι τα καλαθιοφόρα κύτταρα μειώνει την παλίνδρομη καταστολή, που απελευθερώνουν αυτά πάνω στα κοκκιώδη ή στα πυραμοειδή κύτταρα, θέτοντας τα θεμέλια για τη δημιουργία της υπόθεσης των λανθανόντων καλαθιοφόρων κυττάρων (dormant basket cell hypothesis)18. Σύμφωνα με τη θεωρία αυτή, οι ενδιάμεσοι κατασταλτικοί GABAεργικοί νευρώνες του επιληπτικού ιππόκαμπου, λόγω μερικής αποσύνδεσής τους από τις διεγερτικές προσυναπτικές ίνες, βρίσκονται σε "λανθάνουσα", δηλαδή ανενεργό, κατάσταση. Στην περιοχή της οδοντωτής έλικας, αυτή η αποσύνδεση οφείλεται στην απώλεια ενδιάμεσων νευρώνων, οι οποίοι φυσιολογικά θα διεγείρουν τους GABAεργικούς νευρώνες, όπως τα βρυώδη κύτταρα και ενδεχόμενα οι νευρώνες που περιέχουν σωματοστατίνη, ενώ στη CA1 περιοχή η αποσύνδεση των ανασταλτικών κυττάρων αποδίδεται στην απώλεια των πυραμοειδών νευρώνων της CA3 περιοχής στα πλαίσια της ιπποκάμπειας σκλήρυνσης. Η διασπορά των κοκκιωδών κυττάρων συσχετίζεται μ' ένα συγκεκριμένο μοντέλο της εκβλάστησης των παράπλευρων αξονικών κλωνίων των βρυωδών ινών, αλλά η κυτταρική απώλεια του ιππόκαμπου, φαίνεται να είναι το κύριο και καθοριστικό στοιχείο της συναπτικής επανοργάνωσης των βρυωδών ινών στην οδοντωτή έλικα των ασθενών με κροταφική επιληψία21.
Η διατύπωση της θεωρίας των λανθανόντων καλαθιοφορών κυττάρων από τον Sloviter και η πειραματική επαλήθευση της από τους Beckenstein και Lotman έχει σημαντικές θεραπευτικές εφαρμογές, καθώς αποκαλύπτει ότι οι εναπομείναντες πυραμοειδείς νευρώνες του επιληπτικού ιππόκαμπου ανταποκρίνονται στην κατασταλτική δράση του GABA1. Η ενίσχυση αυτής της GABAεργικής καταστολής επιτυγχάνεται με τη χορήγηση της βιγκαμπαντρίνης22. Εντούτοις, σε ό,τι αφορά στην υπόθεση των λανθανόντων καλαθιοφόρων κυττάρων, τα πειραματικά δεδομένα είναι ανεπαρκή για την απόδειξη αυτής. Πρόσφατες καταγραφές από ενδιάμεσους νευρώνες από ιστούς επιληπτικών ασθενών έρχονται σε πλήρη αντίθεση μ' αυτήν23.
Άλλες μελέτες απέδειξαν ότι η μετά από τη χορήγηση καϊνικού οξέος προκαλούμενη βλάβη, περιορίζει την από τα κοκκιώδη κύτταρα της οδοντωτής έλικας απελευθερούμενη καταστολή και προκαλεί μία αναμενόμενη υπερδιεγερσιμότητα. Αυτή η ανώμαλη διεργεσιμότητα αποκαθίσταται και η κατασταλτική δραστηριότητα των κοκκιωδών κυττάρων επανέρχεται όταν ολοκληρωθεί η διαδικασία της εκβλάστησης των παράπλευρων αξονικών κλωνίων των βρυωδών ινών. Αυτή η διαπίστωση οδηγεί στην υπόθεση ότι τα υπό εκβλάστηση παράπλευρα αξονικά κλωνία των κοκκιωδών κυττάρων επανανευρώνουν τους ενδιάμεσους νευρώνες στα πλαίσια μιας προσπάθειας επανορθωτικής διεργασίας (reparative maner). Επιπρόσθετα, αυτά τα δεδομένα υποστηρίζουν την άποψη ότι ο νευρωνικός θάνατος επεμβαίνει άμεσα στην ανάπτυξη της επιληπτικής κατάστασης24.
Παρά το πλήθος των εργασιών, που υποστηρίζουν ότι η ιπποκάμπεια σκλήρυνση οδηγεί στην επιληψία, ακόμη και σήμερα διατηρείται ακέραια η άποψη, ότι οι επιληπτικές κρίσεις παράγουν ή τουλάχιστον επιδεινώνουν την ιπποκάμπεια σκλήρυνση. Πρόσφατη μελέτη καταμέτρησης της νευρωνικής απώλειας στην πορεία του χρόνου, χρησιμοποιώντας τη μέθοδο ανάπτυξης κρίσεων με επαναλαμβανόμενες ηλεκτρικές πυροδοτήσεις (Kindling) αποκάλυψε μία νευρωνική καταστροφή των κυττάρων του ιππόκαμπου, του ενδορρινικού φλοιού και του ουραίου τμήματος του ενδοαπιοειδούς πυρήνα, ανάλογη με τον αριθμό των προκαλουμένων κρίσεων, υποθέτοντας ότι οι επαναλαμβανόμενες επιληπτικές κρίσεις, ίσως είναι η αιτία της ιπποκάμπειας σκλήρυνσης25. Η παρατήρηση σε ανθρώπινο εγκεφαλικό ιστό, ότι οι νευρώνες και σε άλλες περιοχές του εγκεφάλου, όπως, την παρεγκεφαλίδα, τον οπτικό θάλαμο, και τον αμυγδαλοειδή πυρήνα επίσης βλάπτονται στην επιληψία, οδηγεί επίσης στην υπόθεση, ότι οι κρίσεις αυτές καθ' αυτές επηρεάζουν και τελικά καταστρέφουν τα νευρικά κύτταρα8. Η συμβολή των μερικών και των υποκλινικών επιληπτικών κρίσεων στην ιπποκάμπεια καταστροφή είναι άγνωστη και είναι πιθανό μόνο η τροποποίηση της ηλεκτρικής δραστηριότητας σ' αυτό το επιληπτικό υπόστρωμα χωρίς τη συμμετοχή υποξείας, να είναι η μόνη αιτία βλάβης σε ορισμένους νευρώνες. Παρόλα αυτά υπάρχει μία σύγχυση, όσο αφορά στην άποψη ότι οι κρίσεις παράγουν και προάγουν την ιπποκάμπεια σκλήρυνση, καθώς υφίσταται το γεγονός ότι ο επιληπτικός ιππόκαμπος αποκαλύπτει μια ποικιλία νευρωνικής απώλειας χωρίς να εμφανίζει ένα σταθερό τυπικό πρότυπο νευρωνικής βλάβης.
Κλινικά, απεικονιστικά και πειραματικά δεδομένα, υποδηλώνουν ότι η περιοχή του ενδορρινικού φλοιού ίσως να σχετίζεται με την παθοφυσιολογία της ιπποκάμπειας σκλήρυνσης στους ασθενείς με κροταφική επιληψία. Παρόλα αυτά, καμία στατιστικά σημαντική διαφορά δε διαπιστώθηκε στη μέση νευρωνική πυκνότητα στις περιοχές του ενδορρινικού φλοιού μεταξύ των ομάδων των ασθενών με ιπποκάμπεια σκλήρυνση και επιληψία από βλάβες του κροταφικού λοβού26.
Σε πρόσφατες μελέτες, γίνεται προσπάθεια κατάδειξης της ιπποκάμπειας σκλήρυνσης in vivo με τη χρησιμοποίηση της μαγνητικής τομογραφίας (MRI). Όπως φαίνεται, αυτή η νευροαπεικονιστική εξέταση διασαφηνίζει πρώϊμα και πλήρως το σύνδρομο της κροταφικής επιληψίας με ιπποκάμπεια σκλήρυνση. τα χαρακτηριστικά της ιπποκάμπειας σκλήρυνσης είναι η ατροφία του ιππόκαμπου με ένα αυξημένα Τ2 σήμα27.
Μελέτες με τη χρησιμοποίηση MRS (μαγνητική σπεκτροσκοπία), υποστηρίζουν την άποψη ότι η μεταβολική δυσλειτουργία που αξιολογείται με αυτήν, καθώς και η απώλεια του όγκου του ιππόκαμπου που εκτιμάται με την τεχνική της MRI ογκομετρίας, δεν έχουν την ίδια νευροπαθολογική βάση. Τα ευρήματα από την MRS αντανακλούν κυρίως μια νευρωνική και νευρογλοιακή δυσλειτουργία, παρά μια νευρωνική κυτταρική απώλεια28.
Οι ασθενείς που εμφανίζουν κροταφική επιληψία, που σχετίζεται με ιπποκάμπεια σκλήρυνση, κατά κανόνα εκδηλώνουν βαριές διαταραχές της μνήμης και λιγότερο του λόγου και των πρακτικών σχημάτων, οι οποίες φαίνεται να συνδέονται άμεσα με τη βαρύτητα της ιπποκάμπειας σκλήρυνσης αυτής καθαυτής. Έτσι, αυτή η διαπίστωση χρησιμοποιείται στους ασθενείς με κροταφική επιληψία και ιπποκάμπεια σκλήρυνση και με καλή απάντηση στην αντιεπιληπτική θεραπεία σαν δείκτης της εξέλιξης της ιπποκάμπειας σκλήρυνσης29,30.
Συνοψίζοντας το σύνολο των παραπάνω αναφορών, τόσο σε ανθρώπινο όσο και σε πειραματικό υλικό ζώων, συμπεραίνεται, ότι η επιληπτογένεση είναι το αποτέλεσμα της εκλεκτικής απώλειας ορισμένων νευρώνων, οι οποίοι φαίνεται να διαδραματίζουν ένα σημαντικό ρόλο στη μείωση του ουδού για απελευθέρωση της επιληπτικής δραστηριότητας σε ένα κατά τα άλλα φυσιολογικό εγκέφαλο ή σε εγκέφαλο που εμφανίζει μια γενετική προδιάθεση για κρίσεις. Η αιτία για την έναρξη της νευρωνικής απώλειας είναι άγνωστη, αλλά είναι σχεδόν βέβαιο, ότι στα πρώιμα στάδια της επιληπτογόνου διεργασίας, ορισμένοι νευρώνες κλειδιά, εμφανίζουν δυσλειτουργία, η οποία οδηγεί σε επιληπτικές κρίσεις και ιπποκάμπεια σκλήρυνση. Οι επαναλαμβανόμενες κρίσεις με τη σειρά τους, ίσως να επιβαρύνουν τη διεργασία της σκλήρυνσης στον ήδη επιληπτικό εγκέφαλο. ΄Ετσι, μολονότι η μη ειδική κυτταρική απώλεια των νευρώνων του ιππόκαμπου που μπορεί να παρατηρηθεί σε ποικίλες κλινικές οντότητες, δεν σχετίζεται με επιληψία, η ειδική κυτταρική απώλεια ενός νευρωνικού πληθυσμού του ιππόκαμπου, η οποία συνδέεται άμεσα με επιληψία, μπορεί να προκαλεί παθολογική ευερεθιστότητα των διεγερτικών κυκλωμάτων, που οδηγούν σε επαναλαμβανόμενες επιληπτικές κρίσεις.
Στην έρευνα της επιληψίας ένα βασικό ερώτημα είναι ποιοι νευρώνες διαδραματίζουν πρωτεύοντα ρόλο στην έναρξη της επιληπτικής κρίσης. Μολονότι, ελάχιστα είναι γνωστά, γύρω από τους νευρώνες, οι οποίοι χάνονται στην TLE, είναι βέβαιο ότι ο εγκέφαλος δημιουργεί κρίσεις και με την απουσία τους και ως εκ τούτου αυτοί οι νευρώνες δεν είναι απαραίτητοι για την παραγωγή επιληπτικών κρίσεων. Μορφολογική ανάλυση των νευρώνων, που απομένουν στον επιληπτικό κροταφικό λοβό του ανθρώπου, αποκαλύπτει ότι, η πλειότητα αυτών είναι φυσιολογική, αν και ορισμένοι από αυτούς εμφανίζουν μορφολογικές διαταραχές. Μελέτες της μορφολογίας και της κυτταροαρχιτεκτονικής των νευρώνων του επιληπτικού ιππόκαμπου στον άνθρωπο με τον εμπλουτισμό αυτών με άλατα αργύρου (μέθοδος Golgi), αποκαλύπτουν μία απώλεια των δενδριτικών ακανθών σε ορισμένους νευρώνες. Πρόσφατες μελέτες έχουν επίσης αποκαλύψει ένα φάσμα μορφολογικών διαταραχών, που εκτείνεται από μία μείωση έως και πλήρη απώλεια των δενδριτικών ακανθών των πυραμοειδών κυττάρων, μέχρι μια κομβολοειδή διεύρυνση, διόγκωση και παραμόρφωση των κλωνίων του δενδριτικού δικτύου31. Στις περιοχές της γλοίωσης, που πιο συχνά παρατηρείται στην πολύμορφη στιβάδα της πύλης του ιππόκαμπου, τα νευρογλοιακά κύτταρα εμφανίζουν κομβολοειδείς διογκώσεις. Ορισμένα κοκκιώδη κύτταρα της οδοντωτής έλικας, εμφανίζουν απώλεια των δενδριτικών τους ακανθών, κομβολοειδή διόγκωση, καθώς και μείωση της ανάπτυξης του δενδριτικού τους δικτύου. Η σημασία αυτών των μορφολογικών αλλοιώσεων παραμένει άγνωστη. Παρόλα αυτά, οι δενδριτικές άκανθες, όπως είναι γνωστό, είναι θέσεις συναπτικής ανάπτυξης, και ως εκ τούτου, πιστεύεται ότι στον επιληπτικό ιππόκαμπο, η απώλεια των δενδριτικών ακανθών σημαίνει κάποιου τύπου απονεύρωση (denervation). Δεδομένα οδηγούν στην υπόθεση ότι οι δενδριτικές άκανθες, εμφανίζουν μεγάλη πυκνότητα NMDA υποδοχέων, ενώ οι GABAA υποδοχείς είναι αφθονότεροι πάνω στο κυτταρικό σώμα του νευρώνα32, καθώς και στην εκφυτική μοίρα του νευράξονα33. Συμπερασματικά, σ' εκείνους τους νευρώνες που διαπιστώνεται μία μείωση του αριθμού των δενδριτικών τους ακανθών, θα πρέπει να υπάρχει τουλάχιστον μία ανωμαλία στη λειτουργική έκφραση των NMDA υποδοχέων τους.
Πρόσφατα, τα κοκκιώδη κύτταρα της οδοντωτής έλικας, καθώς και οι πολύμορφοι νευρώνες της πύλης του ιππόκαμπου, έχουν γίνει αντικείμενο λεπτομερούς μελέτης. Παρατηρήσεις σε ζώα, αποκάλυψαν ότι, όταν ορισμένα κύτταρα της πύλης καταστραφούν μετά από έγχυση καϊνικού οξέος μέσα στον ιππόκαμπο, οι νευράξονες των κοκκιωδών κυττάρων της οδοντωτής έλικας, οι βρυώδεις ίνες, επανοργανώνονται και δημιουργούν παλίνδρομες συναπτικές σχέσεις μέσα στην εσωτερική μοριώδη (δενδριτική) στιβάδα της οδοντωτής έλικας34,35. Πρόσφατα, παρόμοια επανοργάνωση των βρυωδών ινών παρατηρήθηκε σε ιππόκαμπους επιληπτικών, με τη μέθοδο των επαναλαμβανόμενων ηλεκτρικών πυροδοτήσεων, επιμύων36. Το πρότυπο της επιληψίας από επαναλαμβανόμενη ηλεκτρική πυροδότηση, έχει στενά συσχετιστεί με τη σύνθετη εστιακή επιληψία του ανθρώπου, τόσο από πλευράς κλινικών εκδηλώσεων, όσο και από πλευράς φαρμακευτικής ανταπόκρισης σε ειδικούς αντισπασμογόνους παράγοντες.
Με δεδομένη τη νευρωνική απώλεια στην περιοχή της πύλης στην τυπική ιπποκάμπεια σκλήρυνση, πολλοί ερευνητές μελέτησαν ιππόκαμπους επιληπτικών ασθενών, για να διαπιστώσουν μία πιθανή συναπτική επανοργάνωση στην TLE του ανθρώπου. Ανεξάρτητες μελέτες διαπίστωσαν ότι οι βρυώδεις ίνες, πράγματι δημιουργούν συναπτικές σχέσεις επανοργάνωσης μέσα στην εσωτερική μοριώδη στιβάδα της οδοντωτής έλικας του ιππόκαμπου του ανθρώπου, σχεδόν παρόμοιες με εκείνες που παρατηρούνται σε επιληπτικά πρότυπα ζώων. Για τη διαπίστωση αυτής της συναπτικής επανοργάνωσης, χρησιμοποιήθηκαν δύο τεχνικές: (α) της χρωστικής του Timm με άλατα αργύρου (Timm silver sulfide stain) για βαρέα μέταλλα, η οποία χρωματίζει τις βρυώδεις ίνες, καθώς αυτές περιέχουν υψηλή ποσότητα ψευδαργύρου και (β) της ανοσοϊστοχημικής καθήλωσης της ντινορφίνης Α (dynorphin A), η οποία εμφανίζεται σε υψηλές συγκεντρώσεις, τόσο στα κοκκιώδη κύτταρα, όσο και στα πέρατα των βρυωδών τους ινών. Η επονομαζόμενη εκβλάστηση των παραπλεύρων αξονικών κλωνίων της βρυώδους ίνας (mossy fiber sprouting), περιγράφηκε σε ανθρώπινους εγκεφάλους από τρεις ανεξάρτητες ερευνητικές ομάδες37-39, οι οποίες έδειξαν ότι αυτός ο τύπος εκβλάστησης σχετίζεται με μια κυτταρική απώλεια, τόσο της CA3 περιοχής, όσο και της στιβάδας CA4 της πύλης. Αυτή η διαταραγμένη επανοργάνωση ενοχοποιήθηκε ως βασική αιτία ανάπτυξης της παλίνδρομης ευερεθιστότητας του κοκκιώδους κυττάρου, η οποία φαίνεται να συμμετέχει στους μηχανισμούς της επιληπτογένεσης. Επιπρόσθετα, οι βρυώδεις ίνες δημιουργούν επίσης νέες συναπτικές σχέσεις με όλες τις στιβάδες των CA3, CA2 και CA1 περιοχών και του υποθέματος (subiculum), σε αντίθεση με τη φυσιολογική οργάνωση αυτών στον άνθρωπο, όπου οι βρυώδεις ίνες δημιουργούν συναπτικές σχέσεις μόνο με τη στιβάδα προσανατολισμού (stratum oriens) και τη διαυγή (lucidum) της CA3 περιοχής40. Αυτός ο τελευταίος τρόπος της συναπτικής επανοργάνωσης ενοχοποιήθηκε ότι συμμετέχει σε μια ορθόδρομηπαλίνδρομη διεγερσιμότητα, όχι μόνο στη CA3 περιοχή, αλλά και στο ανατομικό σύμπλεγμα του υποθέματος (subiculum complex). Επιπρόσθετα, θετικά ανοσοαντιδρώντες GABAεργικοί νευράξονες παρατηρήθηκαν σε σχετικά αυξημένη πυκνότητα σε ιστοτεμάχια, που προέρχονταν από ιπποκάμπεια σκλήρυνση, γεγονός που οδηγεί στην υπόθεση ότι και ένας άλλος τύπος εκβλάστησης των παράπλευρων αξονικών κλωνίων από τους νευράξονες των κατασταλτικών ενδιάμεσων νευρώνων εντός των εναπομεινάντων κυτταρικών προσεκβολών, μπορεί να υφίσταται στην TLE του ανθρώπου.
Η μορφολογία των κοκκιωδών κυττάρων, μελετήθηκε σε υλικό που προερχόταν από ασθενείς, οι οποίοι έπασχαν από κροταφική επιληψία με ενδοκυτταρικές εγχύσεις βιοκυτίνης (biocytin) και κίτρινου του Lucifer (Lucifer yellow), oι οποίες σημαίνουν το κυτταρικό σώμα και τις κυτταρικές προσεκβολές του. Δύο κύριοι τύποι ανάπτυξης της προέκτασης των βρυωδών ινών έχουν περιγραφεί. Στον πρώτο τύπο, οι πρωτογενείς άξονες εκτείνονται διαμέσου της πύλης εντός της CA3 περιοχής, απελευθερώνοντας παράπλευρα κλωνία κατά μήκος της διαδρομής τους. Αυτός ο τύπος έντονα προσομοιάζει με τη φυσιολογική ανατομική έκπτυξη των βρυωδών ινών. Ο δεύτερος τύπος έκπτυξης εμφανίζει ένα πρότυπο ανάπτυξης παράπλευρων αξονικών κλωνίων πολύ κοντά στο κυτταρικό σώμα, δημιουργώντας παράπλευρα κλωνία, που εκτρέπονται προς τη στιβάδα των κοκκιωδών κυττάρων και προς τις μοριώδεις στιβάδες (molecular layers). Η ανάπτυξη του δεύτερου τύπου επικοινωνίας, εμφάνιζε εκτεταμένα πλέγματα, τόσο στη στιβάδα των κοκκιωδών κυττάρων, όσο και στην έσω μοριώδη στιβάδα. Χαρακτηριστικό είναι, ότι εκείνοι οι νευρώνες, που παρουσίαζαν εκτρεπόμενα παλίνδρομα αξονικά κλωνία, εμφάνιζαν σημαντικά ογκωδέστερο κυτταρικό σώμα42. Αυτή η μορφολογική διαπίστωση των εκτρεπόμενων αξονικών προβολών, οι οποίες προέρχονται μόνο από τα κοκκιώδη κύτταρα, επιβεβαιώνει τα συμπεράσματα, τα οποία προέρχονται από τα πειράματα με τη χρησιμοποίηση των χρωστικών του Timm και της ντινορφίνης, που εμπλέκουν τα κοκκιώδη κύτταρα της οδοντωτής έλικας στις παθοφυσιολογικές διεργασίες της TLE.
ΒΙΒΛΙΟΓΡΑΦΙΑ
